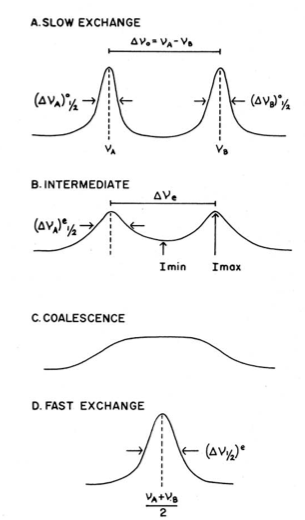
Simulated NMR temperature domains of fluxional molecules. Reprinted with permission from F. P. Gasparro and N. H. Kolodny,
J. Chem. Ed. , 1977, 4, 258. Copyright: American Chemical Society (1977).
Diverse populations
For unequal doublets (for instance, two protons exchanging with one proton), a different treatment is needed. The difference in population can be defined through
[link] , where P
i is the concentration (integration) of species i and X = 2π
Δv t (counts per second). Values for Δ
v t are given in
[link] .
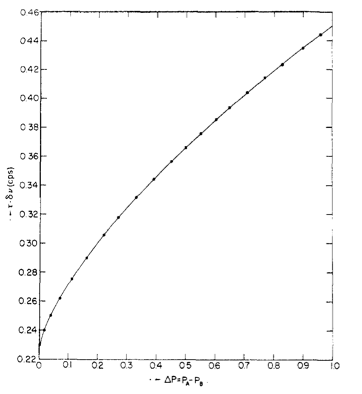
Plot of Δ
v t versus ΔP. Reprinted with permission from H. Shanan-Atidi and K. H. Bar-Eli,
J. Phys. Chem. , 1970,
74 , 961. Copyright: American Chemical Society (1970).
The rates of conversion for the two species, k
a and k
b , follow k
a P
a = k
b P
b (equilibrium), and because k
a = 1/t
a and k
b = 1/t
b , the rate constant follows
[link] .
From Eyring’s expressions, the Gibbs free activation energy for each species can be obtained through
[link] and
[link]
Taking the difference of
[link] and
[link] gives the difference in energy between species a and b (
[link] ).
Converting constants will yield the following activation energies in calories per mole (
[link] and
[link] ).
To obtain the free energys of activation, values of log (X/(2π(1 + ΔP))) need to be plotted against ΔP (values T
c and Δ
v0 are predetermined).
This unequal doublet energetics approximation only gives ΔG
‡ at one temperature, and a more rigorous theoretical treatment is needed to give information about ΔS
‡ and ΔH
‡ .
Example of determination of energetic parameters
Normally ligands such as dipyrido(2,3-a;3′,2′-j)phenazine (dpop’) are tridentate when complexed to transition metal centers. However, dpop’ binds to rhenium in a bidentate manner, with the outer nitrogens alternating in being coordinated and uncoordinated. See
[link] for the structure of Re(CO)
3 (dpop')Cl. This fluxionality results in the exchange of the aromatic protons on the dpop’ ligand, which can be observed via
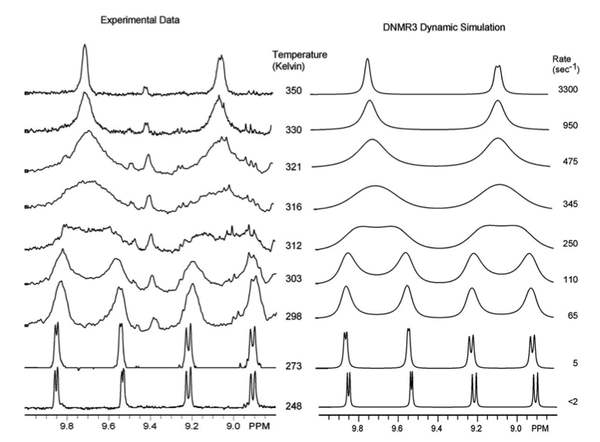
1 HNMR. Because of the complex nature of the coalescence of doublets, the rate constants at different temperatures were determined via computer simulation (DNMR3, a plugin of Topspin). These spectra are shown in
[link] .
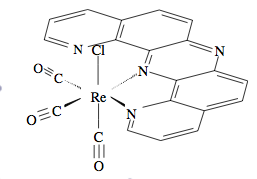
The structure of Re(CO)
3 (dpop’)Cl. Reprinted with permission from K. D. Zimmera, R. Shoemakerb, and R. R. Ruminski,
Inorg. Chim. Acta. , 2006,
5 , 1478. Copyright: Elsevier (2006).experimental and simulated
1 HNMR spectra for Re(CO)
3 (dpop’)Cl. Reprinted with permission from K. D. Zimmera, R. Shoemakerb, and R. R. Ruminski,
Inorg. Chim. Acta. , 2006,
5 , 1478. Copyright: Elsevier (2006).
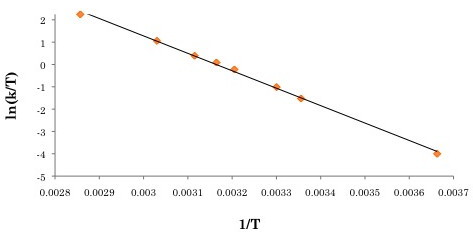
The activation parameters can then be obtained by plotting ln(k/T) versus 1/T (see
[link] for the Eyring plot). ΔS
‡ can be extracted from the y-intercept, and ΔH
‡ can be obtained through the slope of the plot. For this example, ΔH
‡ , ΔS
‡ and ΔG
‡ . were determined to be 64.9 kJ/mol, 7.88 J/mol, and 62.4 kJ/mol.
Eyring plot of ln(k/T) versus 1/T for Re(CO)
3 (dpop’)Cl. Adapted from K. D. Zimmera, R. Shoemakerb, and R. R. Ruminski,
Inorg. Chim. Acta , 2006,
5 , 1478. Copyright: Elsevier (2006).
Limitations to the approach
Though NMR is a powerful technique for determining the energetics of fluxional molecules, it does have one major limitation. If the fluctuation is too rapid for the NMR timescale (<1 ms) or if the conformational change is too slow meaning the coalescence temperature is not observed, the energetics cannot be calculated. In other words, spectra at coalescence and at no exchange need to be observable. One is also limited by the capabilities of the available spectrometer. The energetics of very fast fluxionality (metallocenes, PF
5 , etc) and very slow fluxionality may not be determinable. Also note that this method does not prove any fluxionality or any mechanism thereof; it only gives a value for the activation energy of the process. As a side note, sometimes the coalescence of NMR peaks is not due to fluxionality, but rather temperature-dependent chemical shifts.
Bibliography
H. S. Gutowsky and C. J. Hoffmann,
J. Chem. Phys. , 1951,
19 , 1259.
H. Nakazawa, K. Kawamura, K. Kubo, and K. Miyoshi,
Organometallics , 1999,
18 , 2961.
C. Raynaud, L. Maron, J. P. Daudey, and F. Jolibois.
ChemPhysChem, 2006, 7, 407.
F. P. Gasparro and N. H. Kolodny,
J. Chem. Ed. , 1977,
4 , 258.
B. Kersting, J. R. Telford, M. Meyer, and K. N. Raymond,
J. Am. Chem. Soc. , 1996,
118 , 5712.
M. J. Bennett Jr., F. A. Cotton, A. Davison, J. W. Faller, S. J. Lippard, and S. M. Morehouse,
J. Am. Chem. Soc., 1966,
88 , 4371.
H. Shanan-Atidi and K. H. Bar-Eli,
J. Phys. Chem. , 1970,
74 , 961.
D. J. Press, N. M. R. McNeil, A. Rauk, and T. G. Back,
J. Org. Chem. , 2012,
77 , 9268.
B. D. Nageswara Rao,
Meth. Enzymol. , 1989,
176 , 279.
K. D. Zimmera, R. Shoemakerb, and R. R. Ruminski,
Inorg. Chim. Acta , 2006,
5 , 1478.
Questions & Answers
Discuss the differences between taste and flavor, including how other sensory inputs contribute to our perception of flavor.
The lymphatic system plays several crucial roles in the human body, functioning as a key component of the immune system and contributing to the maintenance of fluid balance. Its main functions include:
1. Immune Response: The lymphatic system produces and transports lymphocytes, which are a type of
asegid
to transport fluids fats proteins and lymphocytes to the blood stream as lymph
Anatomy is the study of the structure of the body, while physiology is the study of the function of the body. Anatomy looks at the body's organs and systems, while physiology looks at how those organs and systems work together to keep the body functioning.
Enzymes are proteins that help speed up chemical reactions in our bodies. Enzymes are essential for digestion, liver function and much more. Too much or too little of a certain enzyme can cause health problems
Kamara
yes
Prince
how does the stomach protect itself from the damaging effects of HCl
the normal temperature is 37°c or 98.6 °Fahrenheit is important for maintaining the homeostasis in the body
the body regular this temperature through the process called thermoregulation which involves brain skin muscle and other organ working together to maintain stable internal temperature
Receive real-time job alerts and never miss the right job again
Source:
OpenStax, Physical methods in chemistry and nano science. OpenStax CNX. May 05, 2015 Download for free at http://legacy.cnx.org/content/col10699/1.21
Google Play and the Google Play logo are trademarks of Google Inc.
Notification Switch
Would you like to follow the 'Physical methods in chemistry and nano science' conversation and receive update notifications?