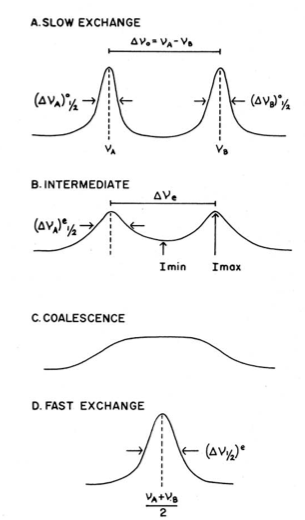
Simulated NMR temperature domains of fluxional molecules. Reprinted with permission from F. P. Gasparro and N. H. Kolodny,
J. Chem. Ed. , 1977, 4, 258. Copyright: American Chemical Society (1977).
Diverse populations
For unequal doublets (for instance, two protons exchanging with one proton), a different treatment is needed. The difference in population can be defined through
[link] , where P
i is the concentration (integration) of species i and X = 2π
Δv t (counts per second). Values for Δ
v t are given in
[link] .
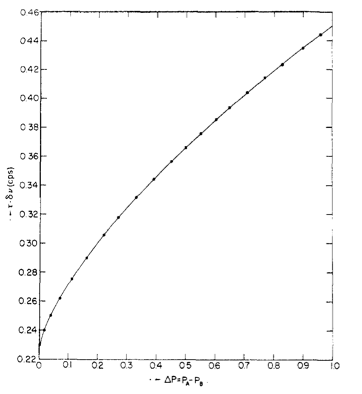
Plot of Δ
v t versus ΔP. Reprinted with permission from H. Shanan-Atidi and K. H. Bar-Eli,
J. Phys. Chem. , 1970,
74 , 961. Copyright: American Chemical Society (1970).
The rates of conversion for the two species, k
a and k
b , follow k
a P
a = k
b P
b (equilibrium), and because k
a = 1/t
a and k
b = 1/t
b , the rate constant follows
[link] .
From Eyring’s expressions, the Gibbs free activation energy for each species can be obtained through
[link] and
[link]
Taking the difference of
[link] and
[link] gives the difference in energy between species a and b (
[link] ).
Converting constants will yield the following activation energies in calories per mole (
[link] and
[link] ).
To obtain the free energys of activation, values of log (X/(2π(1 + ΔP))) need to be plotted against ΔP (values T
c and Δ
v0 are predetermined).
This unequal doublet energetics approximation only gives ΔG
‡ at one temperature, and a more rigorous theoretical treatment is needed to give information about ΔS
‡ and ΔH
‡ .
Example of determination of energetic parameters
Normally ligands such as dipyrido(2,3-a;3′,2′-j)phenazine (dpop’) are tridentate when complexed to transition metal centers. However, dpop’ binds to rhenium in a bidentate manner, with the outer nitrogens alternating in being coordinated and uncoordinated. See
[link] for the structure of Re(CO)
3 (dpop')Cl. This fluxionality results in the exchange of the aromatic protons on the dpop’ ligand, which can be observed via
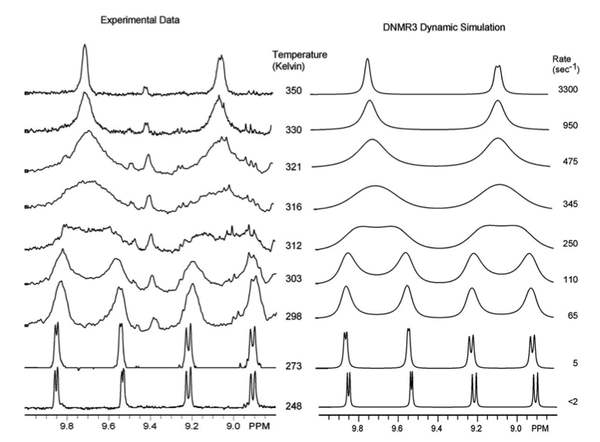
1 HNMR. Because of the complex nature of the coalescence of doublets, the rate constants at different temperatures were determined via computer simulation (DNMR3, a plugin of Topspin). These spectra are shown in
[link] .
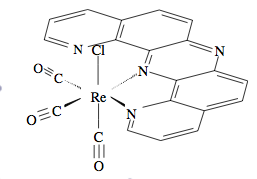
The structure of Re(CO)
3 (dpop’)Cl. Reprinted with permission from K. D. Zimmera, R. Shoemakerb, and R. R. Ruminski,
Inorg. Chim. Acta. , 2006,
5 , 1478. Copyright: Elsevier (2006).experimental and simulated
1 HNMR spectra for Re(CO)
3 (dpop’)Cl. Reprinted with permission from K. D. Zimmera, R. Shoemakerb, and R. R. Ruminski,
Inorg. Chim. Acta. , 2006,
5 , 1478. Copyright: Elsevier (2006).
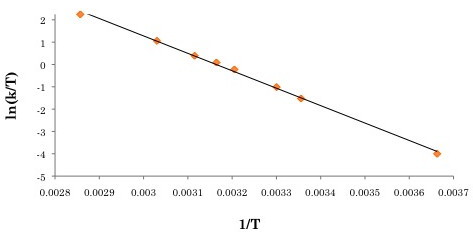
The activation parameters can then be obtained by plotting ln(k/T) versus 1/T (see
[link] for the Eyring plot). ΔS
‡ can be extracted from the y-intercept, and ΔH
‡ can be obtained through the slope of the plot. For this example, ΔH
‡ , ΔS
‡ and ΔG
‡ . were determined to be 64.9 kJ/mol, 7.88 J/mol, and 62.4 kJ/mol.
Eyring plot of ln(k/T) versus 1/T for Re(CO)
3 (dpop’)Cl. Adapted from K. D. Zimmera, R. Shoemakerb, and R. R. Ruminski,
Inorg. Chim. Acta , 2006,
5 , 1478. Copyright: Elsevier (2006).
Limitations to the approach
Though NMR is a powerful technique for determining the energetics of fluxional molecules, it does have one major limitation. If the fluctuation is too rapid for the NMR timescale (<1 ms) or if the conformational change is too slow meaning the coalescence temperature is not observed, the energetics cannot be calculated. In other words, spectra at coalescence and at no exchange need to be observable. One is also limited by the capabilities of the available spectrometer. The energetics of very fast fluxionality (metallocenes, PF
5 , etc) and very slow fluxionality may not be determinable. Also note that this method does not prove any fluxionality or any mechanism thereof; it only gives a value for the activation energy of the process. As a side note, sometimes the coalescence of NMR peaks is not due to fluxionality, but rather temperature-dependent chemical shifts.
Bibliography
H. S. Gutowsky and C. J. Hoffmann,
J. Chem. Phys. , 1951,
19 , 1259.
H. Nakazawa, K. Kawamura, K. Kubo, and K. Miyoshi,
Organometallics , 1999,
18 , 2961.
C. Raynaud, L. Maron, J. P. Daudey, and F. Jolibois.
ChemPhysChem, 2006, 7, 407.
F. P. Gasparro and N. H. Kolodny,
J. Chem. Ed. , 1977,
4 , 258.
B. Kersting, J. R. Telford, M. Meyer, and K. N. Raymond,
J. Am. Chem. Soc. , 1996,
118 , 5712.
M. J. Bennett Jr., F. A. Cotton, A. Davison, J. W. Faller, S. J. Lippard, and S. M. Morehouse,
J. Am. Chem. Soc., 1966,
88 , 4371.
H. Shanan-Atidi and K. H. Bar-Eli,
J. Phys. Chem. , 1970,
74 , 961.
D. J. Press, N. M. R. McNeil, A. Rauk, and T. G. Back,
J. Org. Chem. , 2012,
77 , 9268.
B. D. Nageswara Rao,
Meth. Enzymol. , 1989,
176 , 279.
K. D. Zimmera, R. Shoemakerb, and R. R. Ruminski,
Inorg. Chim. Acta , 2006,
5 , 1478.
Bacteria doesn't produce energy they are dependent upon their substrate in case of lack of nutrients they are able to make spores which helps them to sustain in harsh environments
_Adnan
But not all bacteria make spores, l mean Eukaryotic cells have Mitochondria which acts as powerhouse for them, since bacteria don't have it, what is the substitution for it?
Assimilatory nitrate reduction is a process that occurs in some microorganisms, such as bacteria and archaea, in which nitrate (NO3-) is reduced to nitrite (NO2-), and then further reduced to ammonia (NH3).
Elkana
This process is called assimilatory nitrate reduction because the nitrogen that is produced is incorporated in the cells of microorganisms where it can be used in the synthesis of amino acids and other nitrogen products
There are nothing like emergency disease but there are some common medical emergency which can occur simultaneously like Bleeding,heart attack,Breathing difficulties,severe pain heart stock.Hope you will get my point .Have a nice day ❣️
_Adnan
define infection ,prevention and control
Innocent
I think infection prevention and control is the avoidance of all things we do that gives out break of infections and promotion of health practices that promote life
Receive real-time job alerts and never miss the right job again
Source:
OpenStax, Physical methods in chemistry and nano science. OpenStax CNX. May 05, 2015 Download for free at http://legacy.cnx.org/content/col10699/1.21
Google Play and the Google Play logo are trademarks of Google Inc.
Notification Switch
Would you like to follow the 'Physical methods in chemistry and nano science' conversation and receive update notifications?