

| << Chapter < Page | Chapter >> Page > |
My mentor, Katie Ward, has developed a model to monitor the interactions between the hippocampal cells as well as a number of other types of cells (Grid Cells, Head-direction cells) involved in spatial memory during the Double Rotation experiment. Through collaboration with Dr. James Knierim at Johns Hopkins University, we plan to analyze the interaction of the recurrent connections between CA3 cells in the hippocampus and their influence on the final stability and weight distribution of place fields in order to better understand how these interactions are involved in developing spatial memory.
In order to model spatial memory in the hippocampus, we focus upon the CA3 place cells of the hippocampus. As they have more recurrent connections and show more stability than CA1 cells, they are the most likely candidate for the development of spatial memory.
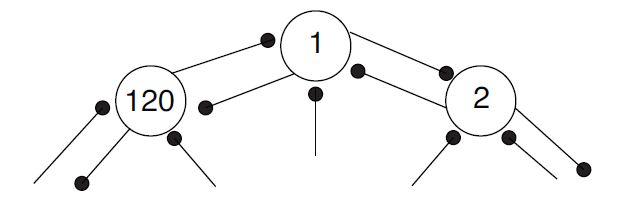
We begin with the experimental setup used by Mehta, where a rat moves clockwise around a circular track. This simplified simulation setup and the parameters given within this chapter were taken from Gabbiani and Cox's book Mathematics for Neuroscientists [link] . In our simplified interpretation of place cell input, each part of the track provides a single place cell with excitatory input of uniform firing rate. We represent the rat's place cell network as a ring of 120 place cells with bidirectional connections between adjacent cells, as depicted in [link] below. As such, each place cell has a three degree window in which it receives external stimulus. If we refer to the figure below and define degree zero as the one at which cell 1 begins to receive stimulus, cell 1 will receive input at degrees 0-3, cell 2 will receive input at degrees 3-6, etc. We denote the cell's place field as the regions on the track during which a cell receives enough stimulus to send signals to neighboring cells. Initially, the cell's place field is synonymous with its three degree window of external stimulus, as it receives enough external input to cause it to stimulate other cells. However, when we introduce synaptic plasticity to our model, we expect the cell's place field to change. We will discuss this in more detail along with synaptic plasticity.
Place cells communicate with each other by transmitting and receiving electrical signals through synapses, just like other neurons. Whether or not the cell sends a signal will depend primarily on its voltage: if it exceeds a specified firing threshold, , the cell will depolarize (reach a positive voltage), send an action potential to neighboring postsynaptic cells, and undergo an obligatory refractory period during which the cell is set to a reset voltage, , and cannot fire again. If the cell voltage does not exceed the threshold, it will remain silent. For our model, we set the threshold to -54 mV and the reset voltage to -60 mV in our simulation. We also use a refractory period = 5 ms.
Notification Switch
Would you like to follow the 'The art of the pfug' conversation and receive update notifications?

|
|

|
|

|
|

|
|
|
|
|

|
|

|
|

|
|

|
|

|